UD 8. CITOLOGÍA: LAS ENVUELTAS CELULARES, EL CITOPLASMA Y EL CENTROSOMA.
BLOQUE II. CITOLOGÍA II. TEMA 8.
LAS ENVUELTAS CELULARES, CITOPLASMA Y CENTROSOMA.


2. La membrana plasmática:
2.1. Composición de la membrana plasmática
2.2. Estructura: Modelo de mosaico fluido
2.3. Funciones de la membrana plasmática
2.4. Uniones intercelulares
2.5. Transporte a través de la membrana plasmática:
- Difusión simple.
- Difusión facilitad.
- Transporte activo
- Transporte de macromoléculas y partículas:
- Endocitosis
- Exocitosis
3. Las membranas de secreción:
- Estructura.
- Funciones del glucocáliz.
- Estructura
- Funciones de la pared celular.
- El citosol.
- El citoesqueleto:
- Componentes del citoesqueleto:
- Microtúbulos
- Microfilamentos o filamentos de actina
- Filamentos intermedios
- Funciones del citoesqueleto.
5. El centrosoma:
- Estructura
- Funciones del centrosoma
CONTENIDOS MÍNIMOS PARA SELECTIVIDAD
Bloque II: “Morfología, estructura y funciones celulares”
3. La célula eucariota: identificar y representar sus partes. Describir la estructura, la función que desempeñan y la relación entre ambas.
3.1. Envueltas celulares. Membrana celular (=membrana plasmática)
* Composición química
* Estructura (modelo mosaico fluido)
* Funciones: Conceptos de transporte pasivo (difusión simple y facilitado) y transporte activo. Procesos de endocitosis y exocitosis.
3.1.1. Funciones de la membrana plasmática:
- Importancia de las membranas en la regulación de los intercambios celulares.
- Transporte de membrana pasivo (difusión simple y facilitada) y transporte activo.
- Endocitosis y exocitosis.
3.2. Envueltas celulares. Pared celular vegetal
3.4. Citoplasma. Conceptos de hialoplasma y citoesqueleto. Centrosoma.
• Composición química. Estructura: modelo del mosaico fluido.
• Los glúcidos como componentes del glicocálix.
• Mecanismos que regulan la fluidez de la membrana. Funciones de la membrana plasmática.


-
INTRODUCCIÓN
Las células presentan una membrana fina y deformable que separa el medio externo de su contenido o citoplasma. Esta membrana está formada por una doble capa lipídica con un elevado número de moléculas de proteínas asociadas. Su función es reconocer su entorno y regular el paso de sustancias.
En el interior celular aparece el citoplasma, en el que se pueden diferenciar: un medio interno o citosol, una red de proteínas filamentosas o citoesqueleto y los orgánulos citoplasmáticos.
En el citoesqueleto destacan los filamentos de actina que bajo la membrana celular edifican un córtex que mantiene la forma celular. También aparecen los filamentos intermedios y los microtúbulos, que organizan y distribuyen los orgánulos citoplasmáticos.

-
LA MEMBRANA PLASMÁTICA
DEFINICIÓN. La membrana plasmática es una lámina fluida, capaz de adoptar cualquier forma, de 75 Å de espesor (6-10 nm - por lo que no pueden verse al microscopio óptico - con el electrónico tienen el aspecto de una doble lámina oscura separada por una banda clara) que separa el interior de la célula (citoplasma) de su entorno y define los diferentes orgánulos del interior de la célula eucariota: retículo endoplasmático, aparato de Golgi, mitocondrias, cloroplastos, lisosomas, etc.
Su característica más crucial es su permeabilidad selectiva, lo que significa que no es una barrera pasiva; al contrario, es la encargada de regular el paso de sustancias que entran y salen de la célula (como nutrientes, iones y desechos), manteniendo el equilibrio interno u homeostasis celular.

La membrana plasmática se comporta como una barrera semipermeable.
2.1. COMPOSICIÓN DE LA MEMBRANA PLASMÁTICA
Todas las membranas biológicas, ya sea la membrana plasmática o la membrana de los orgánulos, tienen una estructura general común: están formadas por una bicapa lipídica fluida semipermeable en la que se incluyen proteínas y glúcidos. Son pues, barreras de permeabilidad selectiva, que controlan las entradas y salidas de sustancias y mantienen unas condiciones estables (homeostasis) en el interior celular.
La composición de la mayoría de las membranas plasmáticas presenta:
-
52% de proteínas,
- 40% de lípidos y un
- 8% de glúcidos.
LÍPIDOS
Los lípidos de la membrana plasmática se encuentran dispuestos en forma de bicapa. Esta bicapa es la estructura básica de todas las membranas biológicas.
Los tres tipos principales de lípidos complejos de membrana son: los fosfolípidos o glicerofosfolípidos (los más abundantes), los esfingoglucolípidos y el colesterol. Dichos lípidos son anfipáticos, es decir, tienen una región hidrófila y otra hidrofóbica; por ello, en un medio acuoso, forman espontáneamente bicapas (autoensamblaje - recuerda micelas) que tienden a cerrarse sobre sí misma (autosellado - ej fecuncación in-vitro), lo que hace posible que los compartimentos formados por las bicapas lipídicas se cierren de nuevo si se rompen.
Enlace vesícula neurotransmisor


Además de estas propiedades de autoensamblaje y autosellado, las bicapas lipídicas tienen otras características importantes que las convierten en una estructura ideal para formar las membranas celulares. Una de las más importantes es la fluidez (propiedad de los cuerpos cuyas moléculas pueden cambiar la posición entre ellas con facilidad)
La bicapa lipídica es fluida (deformable, se adapta al espacio disponible) debido a que las moléculas lipídicas pueden desplazarse libremente.
- El movimiento más frecuente es la difusión lateral dentro de una monocapa;
- Además las moléculas lipídicas giran con gran rapidez alrededor de sus ejes longitudinales mediante rotación
- Y sus cadenas hidrocarbonadas son flexibles, movimiento de flexión.
- Rara vez tiene lugar la difusión transversal de una monocapa a otra, denominada movimiento “flip-flop”.
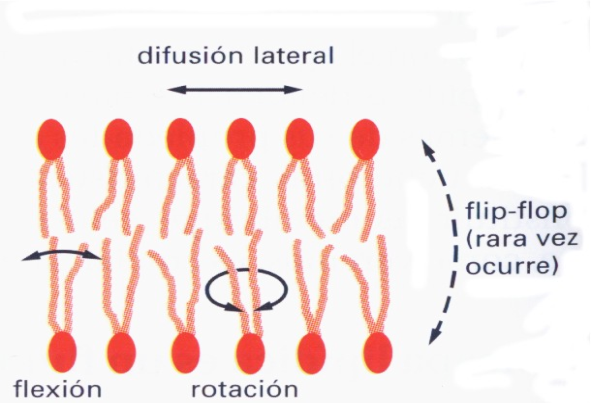
La fluidez de la bicapa lipídica (BL) depende de varios factores:
-
De la temperatura: la fluidez disminuye a medida que desciende la temperatura. La MP solo podrá mantener su fluidez si la temperatura se sitúa por encima del punto de fusión de sus lípidos.
-
De la composición de los ácidos grasos: cuanto más corta sea y más dobles enlaces presenten las cadenas de los ácidos grasos que forman parte de la bicapa lipídica, mayor será la fluidez de la misma.
-
De su contenido en colesterol: el colesterol es el principal regulador de la fluidez de la membrana. El colesterol disminuye la fluidez de la membrana (mb) porque su rígido anillo esteroide interfiere con los movimientos de las cadenas de los ácidos grasos.

COMPETENCIAL 1.. ¿Cuál de los siguientes organismos tendrá una mayor proporcionalidad de ácidos grasos insaturados en sus membranas? Razona tu respuesta.
- Bacterias de manantiales termales.
- Un pez antártico.
- Un ser humano.
- Un cactus.

Otra propiedad de las BL es que, debido a su interior hidrofóbico, son muy impermeables a los iones y a la mayor parte de las moléculas polares. Las moléculas que atraviesan la bicapa son moléculas no polares que se disuelven fácilmente en la bicapa y moléculas polares sin carga de tamaño muy reducido, como el agua, que pasan a través de los huecos que se abren en la bicapa como resultado de los movimientos moleculares. En general, la bicapa actúa de barrera selectiva que controla el paso de sustancias del exterior al interior y viceversa.

PROTEÍNAS
La bicapa lipídica constituye la estructura básica de las membranas biológicas, sin embargo, las proteínas llevan a cabo la mayoría de las funciones específicas de las membranas celulares, por ello, la cantidad y tipo de proteínas de una membrana son un reflejo de su función.
Las principales funciones que desempeñan las proteínas de la membrana son:
-
Estructural: forman parte de la estructura de la membrana plasmática.
-
Transporte: de moléculas específicas hacia el interior o el exterior de la célula.
-
Receptores de las señales químicas del medio y transmitir estas señales al interior celular.
-
Algunas son enzimas que catalizan reacciones asociadas a la membrana (Ej de enzima de MP son las disacaridasas como la lactasa, sacarasa y maltasa en el intestino, que hidrolizan azúcares en monosacáridos).
-
Actuar como puentes estructurales entre el citoesqueleto de la célula y la matriz extracelular desarrollando funciones de reconocimiento y adhesión.
Las proteínas se pueden asociar con la bicapa lipídica de las siguientes formas:
Las proteínas pueden ser:

-
Proteínas transmembrana: atraviesan la bicapa de un extremo a otro. Pueden ser de paso único si una hélice alfa atraviesa la bicapa (1) o de paso múltiple si varias hélices alfa o láminas de una cadena polipeptídica atraviesan la bicapa (2,3). Estas proteínas tienen una parte central hidrofóbica, que interacciona con la región hidrocarbonada de la bicapa y dos partes hidrofílicas, una a cada lado de la bicapa.
-
Proteínas integrales o intrínsecas: se unen mediante enlaces covalentes a los lípidos de la bicapa lipídica. Se localizan en la superficie de la bicapa, ya sea en la cara interna o externa (4,6). No presentan regiones hidrofóbicas.
-
Proteínas periféricas: se unen mediante enlaces no covalentes a otras proteínas (5,7).
Debido a la fluidez de la bicapa, muchas proteínas se desplazan por difusión lateral de forma continua, lo que es esencial para muchas funciones. Sin embargo, otras son prácticamente inmóviles; por ejemplo, determinadas proteínas transmembrana que enlazan el citoesqueleto y la matriz extracelular.
Muchas proteínas son glucoproteínas, ya que llevan oligosacáridos. Se forman en la luz del retículo endoplasmático o del aparato de Golgi, por lo que siempre aparecen en la cara no citosólica de la membrana. Al igual que los lípidos, las proteínas se desplazan por las membranas, pero no pueden invertir su posición: asimetría proteica.
GLÚCIDOS.
Situados en el lado exterior de la membrana, intervienen en funciones de reconocimiento celular. Están unidos a lípidos o a proteínas formando glucolípidos y glucoproteínas.
Los glúcidos constituyen el glucocálix, una membrana de secreción en las células animales formada por dos capas: una, junto a la MP de textura amorfa, y otra, externa, de aspecto fibroso y espesor variable.
Recuerdas

2.2. ESTRUCTURA: MODELO DE MOSAICO FLUIDO (SINGER Y NICOLSON, 1972)
Singer y Nicolson (1972) propusieron el modelo de mosaico fluido para explicar la organización de las membranas biológicas. Según este modelo, las proteínas transmembrana se parecen a icebergs flotando en un mar lipídico bidimensional donde se difunden rápidamente en el plano de la membrana (difusión lateral), a menos que estén ancladas a otros componentes celulares.

Este modelo de mosaico fluido sostiene que:
-
Los lípidos y las proteínas pueden desplazarse en el plano de la bicapa lipídica (difusión lateral)
-
Las membranas son fluidas. La mayor o menor fluidez de la membrana depende de varios factores:
a. A mayor grado de saturación y aumento de la longitud de las cadenas de los ácidos grasos disminuye la fluidez de la membrana.
b. Temperatura: la fluidez disminuye al descender la temperatura.
c. Proporción de colesterol: el colesterol es una molécula plana y rígida, que estabiliza las membranas, es decir, las hace menos flexibles y fluidas.
-
Las membranas son asimétricas, es decir, todas las proteínas de membrana tienen una orientación determinada en la bicapa lipídica, lo que es esencial para su función.
También los lípidos se distribuyen de manera asimétrica. La asimetría es absoluta en el caso de las glucoproteínas y los glucolípidos, ya que los azúcares se localizan siempre en la superficie externa de las membranas plasmáticas.

2.3. FUNCIONES DE LA MEMBRANA PLASMÁTICA
Las funciones principales de la membrana plasmática son las siguientes:
-
La MP limita la célula: separa el citoplasma y sus orgánulos del medio que los rodea.
-
La membrana actúa como una barrera selectiva para el intercambio y el transporte de sustancias entre el interior y el exterior celular.
-
Producción y control de gradientes electroquímicos, ya que en ella se localizan cadenas de transporte y proteínas relacionadas con los mismos (homeostasis)
-
Intercambio de señales entre el medio externo y el medio interno – señales químicas.
-
División celular: la mb está implicada en el control y desarrollo de la división celular y citocinesis.
-
Inmunidad celular: en ella se localizan algunas moléculas con propiedades antigénicas, relacionadas, por ej, con el rechazo de trasplantes de tejidos u órganos de otros individuos, grupos sanguíneos...
-
Endocitosis y exocitosis: la membrana está relacionada con la captación de partículas de gran tamaño (endocitosis) y con la secreción de sustancias al exterior (exocitosis).
-
Mantiene la forma celular, anclando los filamentos del citoesqueleto a los componentes de la matriz extracelular.
2.4. UNIONES INTERCELULARES (No prioritario - Si como función de la mb)

Las uniones de contacto celular son necesarias para construir tejidos. Muchas veces se deben a sustancias intercelulares segregadas por las propias células, las denominadas membranas de secreción, o a especializaciones de la membrana plasmática. Las especializaciones más importantes son:
-
Los Desmosomas son uniones puntuales que dejan un gran espacio intercelular, unos 200 Å. Anclan células, pero sin impedir el paso de sustancias por el espacio intercelular. Estas uniones presentan estructuras proteicas de forma discoidal denominadas placas, de donde parten proteínas transmembrana que se unen fuertemente a las proteínas procedentes de una placa de la célula contigua. El desmosoma se halla unido al citoesqueleto por una red de filamentos de queratina. Los desmosomas se encuentran principalmente en tejidos sometidos a estrés mecánico, como la epidermis (piel), el músculo cardíaco.
-
Las uniones estrechas son uniones que no dejan espacio intercelular. Están formadas por moléculas proteicas transmembrana que se disponen formando hileras que sueldan (cosen) a las membranas plasmáticas entre sí. Estas uniones se hallan reforzadas por proteínas filamentosas intracelulares. No permiten el paso de sustancias, por ello también se denominan uniones herméticas o impermeables. Aparece en los tejidos epiteliales.
-
Las uniones de hendidura o de tipo gap son uniones que dejan un pequeño espacio intercelular. Están constituidas por dos conexones. Un conexón es un fino tubo constituido por seis proteínas transmembranosas que atraviesan la membrana plasmática y se unen a otro conexón de la célula contigua. Son uniones de comunicación, además de anclar células, pone en comunicación sus citoplasmas permitiendo el intercambio de moléculas, por lo que también se les denomina uniones comunicantes. Se encuentran uniendo células del tejido muscular cardíaco.
2.5. TRANSPORTE DE SUSTANCIAS A TRAVÉS DE LA MEMBRANA PLASMÁTICA


EL TRANSPORTE PASIVO
Es un proceso espontáneo, sin gasto energético, de difusión de sustancias a través de la membrana. Se produce siempre a favor de gradiente, es decir, desde el medio donde hay más cantidad hacia el medio donde hay menos, según un gradiente que puede ser:

- Gradiente de concentración. Las moléculas, por simple difusión, pasan desde el medio en donde se encuentran más concentradas hacia el medio en donde su concentración es menor.
- Gradiente eléctrico. Generalmente, el medio celular externo es positivo y el medio celular interno negativo. Por simple difusión, los iones con carga positiva entran en la célula, mientras que los iones negativos salen de ella.
- La conjunción de ambos gradientes origina el gradiente electroquímico, que facilita o reduce la difusión de las moléculas a través de la membrana.
Este transporte puede darse por difusión simple o por difusión facilitada:
- Difusión simple. Se produce a través de la bicapa lipídica. De esta manera atraviesan la membrana plasmática: moléculas no polares liposolubles, como el oxígeno y algunas hormonas lipídicas, como las esteroideas y las tiroideas. También pueden pasar por difusión simple pequeñas moléculas polares sin carga, como el agua, el dióxido de carbono el etanol, el glicerol o la urea.
- Difusión facilitada. Permite el transporte de pequeñas moléculas polares, como los aminoácidos, la glucosa, la sacarosa, o los iones. Estas moléculas al no poder atravesar la bicapa lipídica, requieren la existencia de proteínas transmembranosas específicas para cada sustrato que debe atravesar la membrana. Estas proteínas reciben el nombre de permeasas y canales iónicos.
- 2.1 Proteínas transportadoras o permeasas. Son proteínas transmembranosas que se unen específicamente a la molécula que transportan. Esta unión provoca un cambio de conformación en la proteína que permite que la molécula transportada quede libre al otro lado de la membrana, tras lo cual la proteína transportadora recupera su conformación inicial. Este transporte es específico, ya que cada molécula transportada (azúcares, aminoácidos, metabolitos celulares, etc.) se une en el exterior a un lugar específico de su correspondiente transportador (estado pong), se produce entonces un cambio conformacional (estado ping) por el cual los lugares donde se han unido las moléculas transportadas pasan a estar expuestos al interior celular.

- Proteínas canal o canales iónicos. Son proteínas transmembrana que forman poros acuosos por los que pasan los iones (no experimentan cambios conformacionales). Así, hay canales de Na+, de Ca2+, de Cl-, etc. Además, estos canales están cerrados hasta que reciben una señal adecuada. Estas señales pueden ser químicas (hormonas, neurotransmisores, etc.) en los canales iónicos dependiente de ligando, o eléctricas (cambios en el potencial eléctrico de la membrana) en los canales iónicos dependiente de voltaje.


En la difusión facilitada podemos distinguir dos tipos de transporte:
-
Uniporte: se transporta una única sustancia en un sentido.
-
Cotransporte: se transportan dos sustancias a la vez por la misma proteína transmembrana. Este transporte puede ser de dos tipos:

TRANSPORTE ACTIVO
En este tipo de transporte las moléculas atraviesan la membrana plasmática en contra del gradiente de concentración o del gradiente electroquímico, para lo cual se requiere un aporte de energía. Este proceso de bombeo se realiza mediante proteínas transportadoras denominadas bombas y se consume energía, que se obtiene de la hidrólisis del ATP.
Ejemplos de este tipo de transporte son: la bomba de sodio-potasio, la bomba de calcio y la bomba de protones. En la bomba Na+/K+ la energía derivada de la hidrólisis del ATP se utiliza para expulsar de la célula tres iones de Na+ e introducir dos iones de K+, ambos en contra de su gradiente de concentración. De este modo, contribuye a controlar la presión osmótica intracelular y mantener el potencial de membrana (creando un desequilibrio eléctrico en ambos lados de la membrana. Este sistema es muy importante en la transmisión del impulso eléctrico en las neuronas.). "ver vídeo explicativo"




TRANSPORTE DE MACROMOLÉCULAS Y DE PARTÍCULAS
Endocitosis
En la endocitosis las sustancias que van a ser introducidas en la célula son englobadas en invaginaciones de la MP que acaban cerrándose (autosellado) y formando vesículas intracelulares que contienen el material ingerido. La endocitosis puede ser de 3 tipos:

-
Pinocitosis, cuando la célula ingiere líquidos y sustancias disueltas que almacena en pequeñas vesículas. Ej células o vellosidades del ID.
-
Fagocitosis, cuando la célula ingiere partículas grandes de alimento, o incluso microorganismos (bacterias) y restos celulares mediante la emisión de unas prolongaciones citoplasmáticas denominadas pseudópodos ver vídeo AQUÍ, introduciéndolos en el interior de grandes vesículas denominadas fagosomas. Los materiales fagocitados son digeridos por los lisosomas. La unión de los lisosomas a fagosomas forman los fagolisosomas o vacuolas digestivas, donde se digiere el alimento. (Ej acción de los glóbulos blancos - macrófagos)

Endocitosis dependiente de clatrina. Se utiliza para la entrada selectiva de macromoléculas a las células. En 1º lugar, las macromoléculas que van a ser introducidas quedan englobadas en vesículas en regiones especializadas de la MP llamadas fosas cubiertas, que son pequeñas depresiones de la mb que presentan en su cara citosólica una cubierta formada por la proteína, la clatrina. Un ej de endocitosis dependiente se clatrina es la captación de colesterol - LDL.


Esta imagen representa el proceso de fagocitosis mediada por receptores,(específicamente opsonización) un tipo de endocitosis mediante el cual una célula especializada envuelve y digiere partículas grandes (como una bacteria). Cada número es:
Antígeno / Bacteria: Es la célula extraña o partícula que va a ser ingerida.
Anticuerpos (Opsoninas): Proteínas que se adhieren al antígeno para marcarlo ("opsonización") y facilitar su reconocimiento.
Receptor de membrana: Proteína en la superficie del fagocito que se une específicamente a los anticuerpos o al antígeno.
Pseudópodos: Extensiones de la membrana plasmática y el citoplasma que rodean a la partícula.
Célula fagocítica (Macrófago / Neutrófilo): La célula del sistema inmunitario que realiza la limpieza.
Fagosoma: Vesícula intracelular que contiene la partícula ingerida.
a) Proceso en A e importancia
El proceso señalado con la letra A es la formación de pseudópodos ("falsos pies").
Importancia: Es vital para el sistema inmunitario porque permite a las células de defensa capturar y eliminar patógenos (bacterias, virus) o restos celulares. Sin este mecanismo, las infecciones se propagarían sin control por el organismo.
b) Explicación del proceso (Paso a paso)
Reconocimiento y Opsonización: La bacteria (1) es marcada por anticuerpos (2). Estos sirven como "señal" para que el fagocito la reconozca.
Adhesión: Los receptores (3) de la célula (5) se unen a los anticuerpos que recubren a la bacteria.
Ingestión: La célula emite pseudópodos (4) que se curvan alrededor de la bacteria hasta que sus extremos se fusionan.
Formación del Fagosoma: La bacteria queda completamente encerrada en una vacuola interna llamada fagosoma (6).
Digestión (paso posterior): El fagosoma se unirá a lisosomas (que contienen enzimas) para destruir la bacteria.
Exocitosis
La exocitosis es el proceso contrario a la endocitosis. Mediante este proceso todas las células secretan los materiales necesarios para 1 renovar la MP y los componentes de la 2 matriz extracelular. Igualmente, mediante exocitosis células especializadas en la secreción como las células glandulares vierten al exterior 3 hormonas, neurotransmisores, enzimas digestivas, etc. así como 4 productos de desecho.
Todos los materiales destinados a ser secretados se sintetizan en el retículo endoplasmático, luego pasan al aparato de Golgi donde los productos que van a ser secretados se reúnen y salen en vesículas secretoras que se dirigen a la MP con la que se fusionan para liberar sus contenidos al exterior (hormonas, productos de desecho, moléculas que forman los distintos tipos de matriz extracelulares, etc.)

3. LAS MEMBRANAS DE SECRECIÓN
Son capas constituidas por sustancias producidas por las células que, al ser segregadas, se depositan sobre la superficie externa de la MP. Muchas células animales, que constituyen tejidos, presentan un glucocálix o glucocáliz inmerso en una MB de secreción denominada matriz extracelular, que une las células. En las células vegetales la matriz extracelular se transforma en la pared celular de celulosa.
3.1 Glucocáliz o cubierta celular. (Sólo en células animales)
Se denomina glucocáliz o glicocáliz a la zona periférica de la cara externa de la MP rica en restos glucídicos. Esta cubierta puede tener un espesor de hasta 50 nm, mucho más gruesa que la membrana misma. Es exclusiva de la célula animal.

ESTRUCTURA
El glicocáliz está formado mayoritariamente por las cadenas de oligosacáridos de los glucolípidos y de las glucoproteínas de la MP, aunque también contiene glicoproteínas que han sido segregadas por la célula a la superficie celular.
FUNCIONES
-
Protege la superficie celular del daño mecánico y químico.
Contribuye al reconocimiento y fijación de determinadas sustancias que la célula incorporará mediante fagocitosis o pinocitosis.
Defensa contra el cáncer: los cambios en el glucocálix de las células cancerosas permiten al sistema inmunitario reconocerlas y destruirlas.
-
Reconocimiento celular. La complejidad de los oligosacáridos que forman parte de los glucolípidos y glucoproteínas de la MP, así como el hecho de que se encuentren expuestos en la superficie celular sugiere que estos azúcares son los principales marcadores de identidad que actúa en diversos procesos de reconocimiento celular. Algunos procesos de reconocimiento celular en los que intervienen los azúcares son:
- Procesos infecciosos: Para ejercer su acción dañina las bacterias deben adherirse a la superficie de un tejido y es la unión a los azúcares de la superficie de la célula hospedadora lo que marca el comienzo de la infección.
- Fecundación. Los espermatozoides distinguen las ovocélulas de su propia especie de los gametos femeninos de otras y se unen sólo a los primeros. La especificidad de la fecundación es especialmente importante para los animales acuáticos que vierten sus gametos en el agua, donde pueden mezclarse con gametos de otras especies.
- Reconocimiento entre células de un tejido. Las células que pertenecen a un mismo tejido se reconocen entre sí y son capaces de adherirse y de producir inhibición por contacto en las células vecinas (inhibición de la división celular).
- Rechazo de injertos y trasplantes. Los azúcares de la superficie celular se comportan como antígenos (moléculas que inducen la producción de anticuerpos). Estos antígenos de la superficie celular son característicos de cada individuo y permiten el reconocimiento de las células de un organismo por su sistema inmunitario. La detección de células extrañas por el sistema inmune de un organismo provoca el rechazo de injertos y trasplantes.

LA MATRIZ EXTRACELULAR
En los tejidos el espacio extracelular está ocupado por una compleja red de macromoléculas, denominada matriz extracelular. La matriz está formada por proteínas y polisacáridos muy diversos segregados por las células que están en contacto con ella. Algunas macromoléculas de dicha matriz son luego adsorbidas sobre la superficie celular (por ej, los proteoglucanos (proteína más polisacárido), por ello, es difícil delimitar donde termina la MP y donde empieza la matriz extracelular. Dicha matriz además de unir células y tejidos influye sobre la organización y el comportamiento de las células que rodea.
En los vertebrados, la matriz extracelular del tejido conjuntivo es muy abundante y está formada fundamentalmente por proteínas fibrosas como el colágeno, la elastina, fibronectina y laminina, todas ellas englobadas en un gel de polisacárido hidratado = glucosaminoglucanos (pueden servir de sustancias de reserva en casos extremos) que forman una sustancia gelatinosa. Destaca el ácido hialurónico.

FUNCIONES
-
Mantener unidas a las células, favoreciendo la formación de tejidos.
-
Dar consistencia y elasticidad a los tejidos.
-
Permite la difusión de sustancias.
-
Permite la migración de las células.
-
Rellena los espacios intercelulares.
-
Condiciona la forma de las células.
3.2 La Pared Celular (PC)
La PC de las células vegetales es un tipo especial de matriz extracelular cuya COMPOSICIÓN es:
- Un componente cristalino formado por las fibras de celulosa (quitina en hongos) +
- Y un componente amorfo (sin estructura interna definida) formado por:
- Matriz que une las fibras de celulosa, formada por proteínas, polisacáridos (pectina, hemicelulosa),
- Agua y sales minerales.

-
ESTRUCTURA
La PC puede presentar 3 capas que se forman de manera sucesiva sobre la MP: la lámina media (primera en formarse), la pared primaria y la pared secundaria (última en formarse).
-
Lámina media. Capa más externa. Es compartida por las células contiguas, lo que favorece su unión. Está formada principalmente por proteínas y pectinas que son heteropolisacáridos con carga negativa que tienden a fijar iones de Ca2+ para formar sales insolubles.
-
Pared primaria. Es una capa más gruesa que la lámina media, formada por microfibrillas de celulosa de 5 a 10 nm de diámetro que están dispuestas en planos y a las que se unen moléculas de hemicelulosas, formando haces que se disponen de forma desordenada. Todas estas fibrillas están inmersas en una matriz formada por hemicelulosas, pectinas y proteínas.
-
Pared secundaria. Solamente está presente en algunos tipos de células. Es más gruesa que la pared primaria y es encuentra adosada a la MP. Presenta 3 capas (externa (S1), media (S2) e interna (S3)) en las que las microfibrillas de celulosas se disponen de forma ordenada, paralelos entre sí y dispuestos en planos superpuestos cambiando su orientación al pasar de un plano a otro. Esta pared está formada por celulosa, hemicelulosas y proteínas en menor proporción y no posee pectinas.


Modificaciones/diferenciaciones de la PC:
-
Lignificación: la pared celular se impregna de lignina (polímero muy complejo) para endurecerse. Esto sucede en los vasos leñosos que son células que han perdido las paredes transversales, son tubos por los que sube la savia bruta.
-
Suberificación: la pared celular se impregna de suberina para impermeabilizarla y formar el corcho o súber.
-
Cutinización: la pared se impregna de cutina (ácidos grasos) para impermeabilizar la célula, p.ej en hojas y frutos.
Especializaciones de la PC: la pared primaria y la lámina media son muy permeables al agua, gases y pequeñas moléculas hidrosolubles. El paso de estas sustancias se ve favorecido por la presencia de:
-
Punteaduras: son zonas menos engrosadas de la pared celular, debido a la inhibición del depósito de celulosa y los componentes de la pared celular. También se inhibe el depósito de pared secundaria.
-
Plasmodesmos: son finos conductos o canales que atraviesan las paredes celulares y conectan entre sí los citoplasmas de células adyacentes. Estos canales se forman en la división celular durante la síntesis de la pared, y aparecen en todas las células jóvenes. Los plasmodesmos se encuentran atravesados por retículo endoplasmático que se denomina desmotúbulo y que ayuda a controlar el paso de sustancias a través de los mismos.
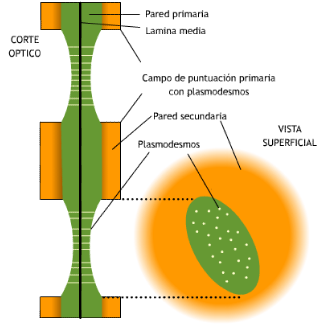
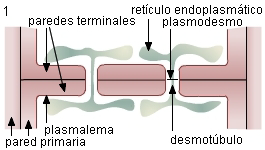
FUNCIONES DE LA PARED CELULAR
La PC desempeña diversas funciones que tienen una enorme importancia para la vida de las plantas. Entre estas funciones hay que destacar las siguientes:
-
Dar soporte mecánico a las células vegetales, actuando como un esqueleto para la planta que permite que se mantenga erguida.
-
Proporcionar la resistencia necesaria frente a los efectos de la ósmosis.
-
Proteger frente a la abrasión mecánica y frente al ataque de insectos y de microorganismos patógenos.
-
Participar en la comunicación entre células, principalmente a través de los plasmodesmos, por los que pasan distintos tipos de moléculas de pequeño tamaño.
-
Orientar el crecimiento de células y tejidos y participar en la diferenciación celular.
La PC comienza a formarse como una lámina fina que aparece entre las MP de las dos células resultantes de la división celular. Su desarrollo posterior se hace mediante la síntesis de sus componentes: las proteínas, hemicelulosas y pectinas es sintetizada por los orgánulos celulares, sin embargo la celulosa es sintetizada en la MP por un complejo enzimático denominado Celulosa sintasa.
Recuerda la composición de otra paredes celulares.
- Bacterias. Peptidoglucano (arqueas carecen de ello)
- Hongos y exoesqueleto de artrópodos. Quitina o N-Acetilglucosamina.
- Algas y plantas. Celulosa. Homopolisacárido de glucosa beta.
- Diatomeas y radiolarios. Silicio.
4. EL CITOPLASMA

4.1. El citosol o hialoplasma
El citosol, también llamado citoplasma fundamental o hialoplasma, se encuentra delimitado por el sistema membranoso celular, es decir, ocupa el espacio situado entre la membrana plasmática, la envoltura nuclear y las membranas de los diferentes orgánulos. En él están inmersos el citoesqueleto, los ribosomas y los demás orgánulos.
Nota - citosol = disolución sin orgánulos ni núcleo / Citoplasma incluye orgánulos (pero no el núcleo) y citoesqueleto
ESTRUCTURA DEL CITOSOL
Es un medio acuoso con un 85% de agua, en el cual aparecen disueltas una gran cantidad de moléculas formando una dispersión coloidal. Estas moléculas son: proteínas, aminoácidos, enzimas, lípidos, glúcidos, ácidos nucleicos (ARNt y ARNm), nucleótidos (como el ATP), nucleósidos, productos del metabolismo y sales minerales disueltas. El citosol puede presentar dos estados físicos: el estado gel (viscoso) y el estado sol (fluido).

FUNCIONES DEL CITOSOL
Las funciones del citosol son las siguientes:
-
Regulador del pH intracelular.
-
Lugar donde se almacenan sustancias - Inclusiones citoplasmáticas = Son materiales almacenados en el citoplasma y no rodeados de membranas. Destacan el glucógeno y las grasas (reservas de energía). El Glucógeno, al microscopio electrónico aparece como gránulos dispersos, sobre todo en células del hígado y músculo. Recuerdas: Grasas son la fuente de energía más importante. Proporciona el doble de energía por gramo que los glúcidos y, además, ocupa menor espacio. La mayor parte de las grasas se almacenan en el citoplasma del tejido adiposo, como una gran gota de triglicéridos.
En él tienen lugar movimientos de ciclosis. corrientes citoplasmáticas rotatorias cuya función es la de facilitar el intercambio de sustancias intracelularmente o entre la célula y el exterior.
-
Lugar donde se realizan múltiples reacciones metabólicas:
- Gluconeogénesis.
- Glucólisis.
- Biosíntesis de aminoácidos.
- Biosíntesis de ácidos grasos.
- Síntesis de proteínas.
- Modificación de proteínas
4.2. El citoesqueleto
El citoesqueleto es exclusivo de las células eucariotas y está formado por tres tipos de filamentos: los microtúbulos, los microfilamentos o filamentos de actina y los filamentos intermedios, que interaccionan entre sí y se unen a los orgánulos celulares y a la membrana plasmática mediante un conjunto de proteínas accesorias.
El citoesqueleto de las células es una estructura dinámica que se reorganiza continuamente según las células se mueven o cambian de forma y durante la división celular.


PROPIEDADES DE LOS COMPONENTES DEL CITOESQUELETO
Los microfilamentos o filamentos de actina y los microtúbulos:
-
Son estructuras polares, es decir, que sus dos extremos tienen propiedades distintas: uno de ellos crece a gran velocidad uniendo monómeros de actina (microfilamentos) o tubulina (microtúbulos), mientras que el otro crece lentamente. El extremo de crecimiento rápido se denomina extremo más (+) y el otro extremo menos (-).
-
Son estructuras lábiles, es decir, pasan por fases de crecimiento (por adicción de monómeros en los extremos) y de acortamiento (por pérdida de monómeros de los extremos).
-
La estabilidad de los filamentos de actina y de los microtúbulos está regulada por las proteínas asociadas.
Los filamentos intermedios:
A diferencia de microfilamentos y microtúbulos, son estables, no polares y las proteínas fibrosas que los forman varían de unas células a otras, por lo que los filamentos intermedios reciben nombres distintos según el tipo de célula que se trate, por ejemplo, en el sistema nervioso se denominan neurofilamentos.
5. EL CENTROSOMA O CITOCENTRO
El centrosoma o centro celular es una estructura sin membrana, presente en todas las células animales susceptibles de dividirse. Salvo algunas excepciones, no existe en células vegetales.
ESTRUCTURA
El centrosoma consta de un cuerpo central formado por dos centriolos, denominados diplosoma, que se encuentran rodeados por el material periocentriolar denso y amorfo con cientos de estructuras en forma de anillos, compuestas por un tipo especial de tubulina que sirven como punto de nucleación para la formación de microtúbulos. Los microtúbulos se unen a estos anillos por sus extremos menos (-), que quedan anclados, y el crecimiento tiene lugar hacia sus extremos más (+) que se alejan del centrosoma. Los microtúbulos que parten del centrosoma se denominan Áster. En las células animales, el centrosoma se sitúa cerca del núcleo y desde ahí irradian los microtúbulos extendiéndose por el citoplasma.

Ambos centriolos del diplosoma se disponen perpendicularmente entre sí, presentando una estructura cilíndrica de 0,2 µm de diámetro, cuyas paredes están formadas por nueve grupos de tres microtúbulos o tripletes que forman la denominada estructura 9 x 3 + 0.
Los tres microtúbulos que constituyen cada triplete están estrechamente asociados los unos a los otros, y ligeramente desplazados. Se denominan microtúbulo A (el más interno y más próximo al eje del cilindro), microtúbulo C (el más externo) y microtúbulo B (situado entre los anteriores). Los tripletes que se encuentran adyacentes están unidos entre sí mediante una proteína denominada nexina.

El microtúbulo A es completo (formado por 13 protofilamentos); sin embargo, los microtúbulos B y C son incompletos, están formados por tan sólo diez protofilamentos. Cada uno de estos microtúbulos comparte tres protofilamentos con el anterior, el A o el B, respectivamente.
En el centriolo se distinguen un extremo proximal cercano al núcleo celular, y un extremo distal dirigido hacia la periferia. En el interior de la región proximal se observa un material denso del que salen una fibrillas radiales dirigidas hacia la cara interna de los microtúbulos A. Esta estructura proporciona una imagen conocida como estructura en rueda de carro.


FUNCIONES
Las funciones más importantes del centrosoma son las siguientes:
-
Es el centro organizador de microtúbulos de la célula.
-
De él derivan las estructuras constituidas por microtúbulos como los cilios y flagelos, encargados del movimiento celular.
-
Origina el huso acromático encargado de la separación de los cromosomas durante la división celular
-
Controla la estructura del citoesqueleto, cuyos filamentos se organizan alrededor de los microtúbulos.
Las células vegetales sin centrosomas construyen sus microtúbulos a partir de un centro organizador de microtúbulos, similar al centrosoma de las células animales.
COMPETENCIALES.
1. La figura representa esquemáticamente un fragmento de membrana plasmática.
-
Indica las características moleculares de los elementos señalados por los números 1 a 3.
-
Indica una posible función de los elementos señalados con el número 3 y dos posibles funciones de los elementos señalados con el número 2.
-
Las células poseen numerosas membranas internas que constituyen una parte importante de sus orgánulos. La composición y propiedades de esas membranas no es la misma en todos los casos. Indica dos ejemplos de estas diferencias.
¿Qué tipos de lípidos están presentes en las membranas celulares?
¿Qué diferencia hay entre proteína integral y proteína periférica?
¿Qué significa el término de modelo mosaico fluido?
Cita 3 orgánulos delimitados por membranas.
3. La membrana plasmática no es una estructura estática, ya que los lípidos que la componen tienen posibilidad de moverse dentro de ella, lo que le proporciona una cierta fluidez o viscosidad. Explica detalladamente los movimientos que pueden realizar los lípidos y la importancia que el colesterol tiene para la funcionalidad de la membrana plasmática.
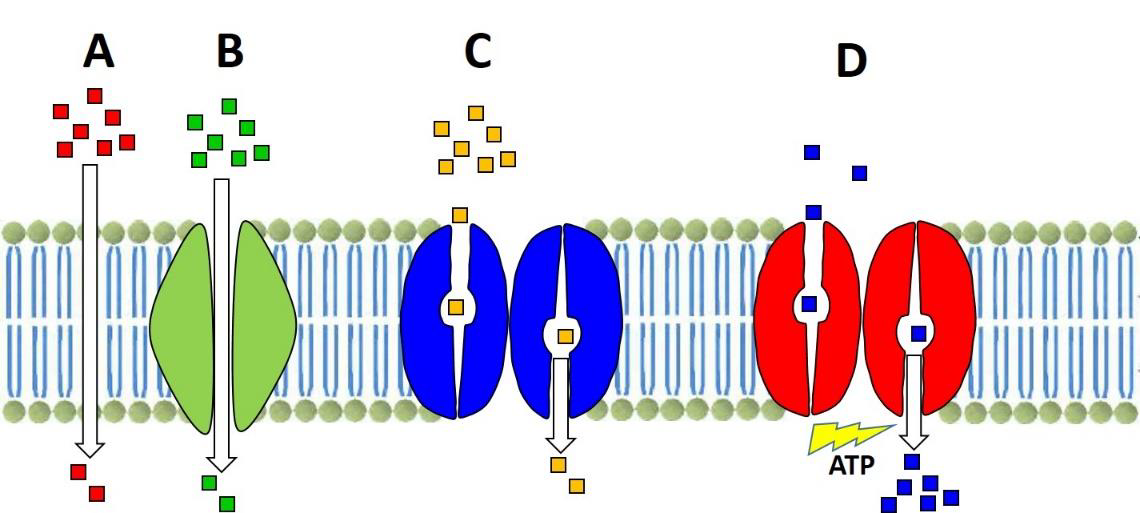
a. Identifica los tipos de transporte representados por las letras A, B, C y D (1 punto)
b. Señala qué condiciones debe tener una molécula para que pueda pasar mediante el transporte indicado con la letra A. Indica dos ejemplos de moléculas que pasen así. (1P)
c. ¿Cómo se llama el complejo proteico de transporte que permite el paso de ARN mensajero desde el núcleo al citoplasma? Indica dos de sus características principales. (0,5 puntos)
5. La figura esquematiza un fragmento de membrana plasmática. Responde a las siguientes cuestiones:
-
Indica las características moleculares de los elementos señalados con números.
-
Indica dos posibles funciones de los elementos señalados con el número 2.
-
Indica, muy brevemente, qué son los plasmodesmos y los desmosomas.
6. En el siguiente esquema se muestra una pareja de centriolos, Eª también conocida como diplosoma:
-
¿Qué tipo de células presentan esas estructuras?
-
¿Qué papel juega en la división celular?
-
¿Qué relación presentan estas estructuras con los cilios y flagelos?
-
La colchicina es una sustancia que despolimeriza los microtúbulos. ¿Qué consecuencias tendría para una célula en división la acción de la colchicina?
7. Indica los componentes, y describe la organización y funciones de la pared celular en c vegetales.
8. Respecto a la membrana plasmática de las células eucariotas animales:
A. Indique dos características del modelo de mosaico fluido (0,5 puntos)
B. Explique la acción de dos factores que aumenten la fluidez de la membrana (0,5 puntos)
C. Si la célula es vegetal, ¿puede llevar esteroles en su membrana? Razone la respuesta (0,4 p)
D. Indique dos diferencias existentes entre transporte pasivo y transporte activo a través de la membrana plasmática (0,4 puntos). Ponga un ejemplo de cada caso (0,2 puntos)
9. Se está llevando a cabo un estudio en células animales para determinar el efecto de un medicamento sobre las membranas celulares. Al aplicar dicho medicamento se observa que el número de dobles enlaces presentes en los ácidos grasos aumenta considerablemente respecto al contenido de las células control.
a) Indique la estructura básica de una molécula de ácido graso [0,25].
b) Explique razonadamente qué efecto puede tener sobre la membrana celular este incremento de dobles enlaces [0,75].
c) Defina membrana plasmática e indique una función de esta estructura [0,5]. d) Nombre dos orgánulos celulares delimitados por una membrana simple e indique una función que desempeñen [0,5].
10. Describa el modelo de mosaico fluido de membrana, y cita al menos 3 de sus funciones. Represéntelo con un dibujo en el que aparezcan claramente señalados los distintos tipos de lípidos, proteínas y glúcidos que la conforman.
PAU 25. COMPETENCIAL +-
PAU25 ZARAGOZA.. (CON SOLUC)
4. Elija únicamente una de las dos opciones que se plantean:

Opción 4.A) A continuación se visualiza la siguiente estructura de una célula animal. (2 puntos)
a) ¿Qué estructura representa la siguiente imagen? Justifíquelo (0,3 puntos)
b) Identifique las estructuras señaladas con los números 1 al 7. (0,7 puntos)
c) Si esta estructura presentara una gran cantidad de moléculas identificadas con el número 2, ¿qué le
ocurriría a esta estructura? Justifique su respuesta (0,2 puntos)
d) ¿En qué se parecen y en qué se diferencian las estructuras A y B en cuanto a su función? (0,8
puntos)
PAU25 CANARIAS. Bloque III: Se debe seleccionar 2 preguntas

11. La apitoxina es el veneno secretado por las abejas. Cuando el aguijón de una abeja obrera es introducido en la piel, unos 0,1 a 0,3 mg de esta sustancia es inyectada con un efecto citotóxico, provocando la destrucción de las membranas celulares.
a. La figura corresponde a la estructura vital que es la membrana plasmática, identifica los componentes enumerados en el esquema.
b. Con algún componente de la figura relaciona las funciones siguientes: el reconocimiento celular, la fluidez y el transporte de moléculas.
c. ¿Qué significa el término de mosaico de fluido?
d. Ciertas células eucariotas presentan otra estructura externa, además de la representada en el esquema. ¿A qué estructura nos referimos?
PAU25. ASTURIAS.
11. En relación al transporte a través de la membrana:
a. ¿En qué se diferencian el transporte activo y el pasivo?
b. ¿ A qué tipo de transporte corresponde la difusión simple?
c. ¿Qué clase de moléculas atraviesan la membrana plasmática mediante el transporte por difusión simple?
d. ¿En qué se diferencian los mecanismos de pinocitosis y fagocitosis?
PAU25 CASTILLA Y LEÓN.Pregunta 3.1. Biología celular (2 opciones a elegir 1) (1,5 puntos)
Opción 3.1.A. Conteste a las siguientes cuestiones sobre la organización celular:
a) Cite una función de cada uno de los siguientes orgánulos: lisosomas, retículo endoplasmático liso, aparato de Golgi y nucléolo. (0,8)
b)Señale dos diferencias entre el transporte pasivo y el transporte activo, y cite un ejemplo para cada uno de ellos.
PAU25. CANTÁBRIA.
Pregunta 3. El esquema representa la membrana plasmática de una célula eucariota animal.

Opción A
1. Indica cuatro tipos de moléculas que formen parte de la membrana plasmática y que se representen en el esquema. Indica cuál de ellos participa en el reconocimiento celular. (Calificación 1 punto)
2. Indica cómo se llama el movimiento de un componente de membrana de una hemimembrana a otra, señala cuál es el compuesto que con mayor facilidad puede hacer este tipo de movimiento, explica por qué y señala qué función desempeña este compuesto en la membrana. (Calificación 1 punto)
3. Indica si, en la membrana que se representa en el esquema, la hemimembrana externa y la hemimembrana interna tendrían el mismo o diferente grado de fluidez. Justifica tu respuesta. (Calificación 0.5 puntos)
Opción B
1. Indica el nombre del mecanismo de transporte que se representa en la figura y señala en qué dirección tendrá lugar el transporte de H2O si la presión osmótica es más alta en el lado extracelular que en el lado intracelular de la membrana. (Calificación 1 punto)
2. Indica cómo se llama el mecanismo que permite transportar de forma específica hacia el interior de la célula macromoléculas de elevada masa molecular. Explica brevemente cómo tiene lugar este tipo de transporte. (Calificación 1 punto)
3. Explica qué es el potencial de membrana y señala qué elemento de la membrana es esencial para generar este potencial y cómo lo hace. (Calificación 0.5
EJERCICIO 3
(2 preguntas, de las que debe responder, a su elección, SOLAMENTE UNA)
Pregunta 3.1 (2 puntos)
En relación con la figura adjunta:

a) ¿Qué estructura representa la imagen? [0,2].
b) Identifique los componentes señalados con las letras A, B, C, D, E, F y G [0,7].
c) Relacione las siguientes funciones con algún componente celular de los identificados: fluidez, reconocimiento celular,transporte de sustancias [0,6].
d) Explique la estructura y composición del componente señalado con la letra D, relacionándolo con su función [0,5].
PAU25. NAVARRA.
6. El transporte de glucosa es esencial para la supervivencia y función de las células humanas.
a) Indica los diferentes mecanismos de transporte a través de la membrana celular y las principales características. (0.75P)
b) De estos mecanismos, ¿cuáles podría utilizar la glucosa para transportarse en las células humanas? (0.25 P)
PAU25. MADRID
3c) En un experimento se van a utilizar tres vesículas artificiales con distinta composición en su membrana:
Vesícula A: con ácidos grasos saturados y sin colesterol.
Vesícula B: con ácidos grasos insaturados y sin colesterol.
Vesícula C: con ácidos grasos saturados y con colesterol.
Justifique cuál de ellas es la más fluida. Justifique cuál será la más apta para realizar la endocitosis y la exocitosis (0,75 puntos).
4.-B.- En relación con las membranas biológicas:
a) Relacione cada característica de la columna de la izquierda con un único concepto de entre los de la derecha (no hace falta que
copie el texto, solo que empareje los números y letras que identifican cada opción) (1 punto).
2. Un catión pasa por una proteína canal B. Difusión facilitada
3. Célula vegetal en medio hipotónico C. Simporte activo secundario
4. Entrada de O2 gas D. Antiporte activo secundario
5. La glucosa entra a través de una permeasa E. Transporte activo primario (bomba)
6. Entran iones contra gradiente electroquímico
con gasto de ATP F. Turgencia
7. Entra un aminoácido contra gradiente junto
con un ion a favor de gradiente G. Difusión simple a través de la bicapa lipídica
8. Entra un aminoácido contra gradiente y sale
un ion a favor de gradiente. H. Plasmolisis
b) Indique dos funciones de las membranas distintas de la permeabilidad selectiva y el transporte de compuestos (0,5 puntos).
c) Indique los tres componentes principales de la membrana plasmática y describa brevemente su localización en la misma (0,5puntos).
PAU25 VALENCIA. b) ¿Cuál es la estructura que se observa en la imagen? Explique brevemente su funcionamiento (1 unto).
CITOESQUELETO.
PAU25. MURCIA. En un periódico se podía leer la siguiente noticia: El Grupo de Síntesis Orgánica de la Universitat Jaume I ha desarrollado un derivado de la colchicina, uno de los fármacos contra el cáncer más potentes. Tal y como explican los investigadores, “la colchicina es uno de los compuestos naturales con mayor capacidad para detener la reproducción de las células cancerosas, pero su elevada toxicidad impide que pueda ser utilizado en tratamientos extensos como los que requieren los pacientes oncológicos". Fuente: El Mundo 5 oct 2018
Para explicar la noticia a su padre, ha buscado y encontrado la siguiente información: “La colchicina es una sustancia química altamente tóxica para ciertas células; es un compuesto que bloquea la mitosis e interrumpe el transporte a lo largo de los axones de las neuronas. La colchicina inhibe la dinámica de formación de los microtúbulos uniéndose a la tubulina”.
A) Teniendo en cuenta esta información y sus conocimientos sobre la composición y la función de los microtúbulos, explique cómo interfiere esta droga en: i) la reproducción de las células (0,5 puntos); ii) el correcto funcionamiento de las neuronas (0,5 puntos).
B) Nombre otro componente del citoesqueleto de células eucariotas e indique su composición y una de sus funciones (0,5 puntos).
PAU25 ASTURIAS. Pregunta 3. El esquema adjunto es una representación esquemática de una célula y el recuadro de la derecha corresponde detalle a mayor aumento de una estructura presente en el citoplasma.

10. La microfotografía y el esquema que la acompaña corresponden a un corte transversal de una determinada prolongación celular.
a. ¿De qué estructura se trata?
b. ¿Cuál es su naturaleza química?
c. ¿En qué función biológica está implicada?
d. ¿Qué 3 tipos de elementos pueden formar parte del citoesqueleto?

EBAU - NO COMPETENCIALES.
1. EBAU 2018 y Ej3. Estructura, composición y tamaño de la membrana plasmática: Modelo del mosaico fluido. (2p)
2. EBAU. Respecto a la membrana plasmática:
A. Factores que determinan la fluidez de la membrana plasmática (1p)
B. Describe 5 funciones de las membranas plasmáticas. (1p)
3. EBAU. 4.- Permeabilidad de la membrana plasmática.
A) Diferencias entre transporte activo y pasivo a través de la membrana celular. (0,5 p)
B) Enumere y explique los diferentes tipos de transporte pasivo a través de la membrana celular. (1,5 p)
5. EBAU 2019 20. 3.- Respecto de la membrana plasmática:
A. Indique cuatro componentes de la membrana plasmática de una célula animal. (0.5 puntos)
B. Diferencie transporte pasivo y transporte activo. (0.5 puntos)
C. Explique el transporte pasivo. (1 punto)
B. Dos funciones de la pared celular (0.5 puntos).
EBAU 2018 4. Diferencias estructurales entre la pared celular vegetal y la pared bacteriana. (2p)


Comentarios
Publicar un comentario